Adrenergični nervni sistem i zatajenje srca. Adrenergični nervni sistem
ADRENERGIČNI LIJEKOVI
ADRENOMYMETICS
a1 a2 b1 b2 Adrenalin hidrohlorida1 a2 b1 Norepinefrin hidrotartrat
a1 Mezaton
a2 Klonidin = Klonidin
Guanfacine = Estulic
Naphthyzin
Galazolinv1 v2 Izadrin
Orciprenalin sulfat = Alupent
u 1 Dobutamin
u 2 Fenoterol = Berotec = Partusisten
Formoterol
Salmeterol
Salbutamol
Terbutalin
Klenbuterol=Kontraspasmin
SYMPATOMIMETICS
FenaminADRENO BLOCKERS
α-blokatori
a1 a2 neselektivni
fentolamin
piroksan
Dihidrogenirani alkaloidi ergot
α 1 adrenolitici
Pra zozin=Patsiol
Doxa zozin=Tonocardin
Tera zozin=Cornam
β-blokatori
Kardioselektivni
Talino lol=Cordanum
Ateno lol=Tenormin
Metopro lol=Betalok
Alcebuto lol=Sektralno
Betaxo lol=Lokren
Bisopro lol= Concor
Cardionon-selektivni
Proprano lol=Anaprilin
Oxpreno lol=Trazicor
Pindo lol=Whisken
Saće lol
Sa VSA "intrinzičnom simpatomimetičkom aktivnošću"
Oxpreno lol
Atsebuto lol
αβ-blokatori
Labeta lol
Karvedi lol
SYMPATHOLYTICS
Methyldopa=Dopegyt=AldometOctadine=Guanetidin=Izobarin
Ornid=Bretylium tosylate
Reserpine=Rausedil
PRENOS U ADRENERGIČKIM SINAPSIMA
STRUKTURA I FUNKCIJA SINAPSA
Synapse– funkcionalni (hemijski) kontaktDvije nervne ćelije ili
Nervne ćelije i ćelije izvršnih organa
U sinapsama postoje 2 membrane:
presinaptička membrana akson -
odašiljanje
Postsinaptička membrana nervna ćelija ili ćelija izvršnog organa – perceptivna
Sinaptički rascjep
Nalazi se između membrana
Napunjen polisaharidnim gelom
Ima pore za difuziju medijatora
Ograničeni elementima vezivnog tkiva (sprečavaju oslobađanje medijatora u krv)
sinaptičke vezikule - depo medijatora (u vezi sa proteinima)
Tokom odmora potencijal pojedinačni dijelovi odašiljača se oslobađaju u sinaptički pukotinu -
za održavanje fizioloških reakcija organa i tonusa skeletnih mišića
Tokom akcionog potencijala
Pozitivni naboj na unutrašnjoj površini presinaptičke membrane uzrokuje da se negativno nabijene sinaptičke vezikule zalijepe za nju.
Kalcijumovi joni katalizuju interakciju presinaptičkih membranskih proteina sa proteinima sinaptičkih vezikula.
U presinaptičkoj membrani se otvara kanal za oslobađanje dijela transmitera u sinaptičku pukotinu.
Nakon interakcije sa receptorom
transmiteri nestaju iz sinaptičke pukotine kao rezultat:
Neuronsko uvlačenje
(povratak u sinaptičke vezikule da učestvuje u retransmisiji impulsa)
Ekstraneuronsko uvlačenje
(izlaganje u izvršnim organima)
Enzimska probava
PRENOS U ADRENERGIČKIM SINAPSIMA
koji se provodi korištenjem kateholamina
norepinefrin -glavni posrednik
dopamin- rjeđe igra ulogu posrednika
adrenalin- ćelije medule nadbubrežne žlezde proizvode i
pusti u krv, tj. to je hormon
Postojanje tri kateholaminska medijatoraevolucijski određeno, a ne slučajno. Svaki od njih ima afinitet za određenu vrstu receptora, zbog čega nervni sistem može višediferenciranutiču na funkcije organa.
Organi sa simpatičkom inervacijom
Gotovo sve sadržano u njima norepinefrin lokalizovan u nervnim vlaknima.
U ćelijama srži nadbubrežne žlijezde kateholamini nalazi u hromafinskim granulama.
Postoje dvije vrste ćelija koje sadrže kateholamine u meduli nadbubrežne žlijezde
-sa norepinefrinom
- sa adrenalinom.(U ovim ćelijama norepinefrin ostavlja hromafinske granule u citoplazmu,
ovdje je metiliran u adrenalin.
Adrenalin ponovo ulazi u granule i tamo se čuva dok se ne oslobodi.
Kod odraslih, adrenalin čini 80% svih kateholamina
moždana materija, 20% - norepinefrin.)
Glavni faktor koji regulira brzinu sinteze adrenalina su glukokortikoidi.
Glukokortikoidi ulaze u sistem portala nadbubrežne žlijezde.
Dugotrajni stres, koji uzrokuje pojačano lučenje ACTH,
dovodi do povećanja sinteze hormona i kortikala (kortizola),
i medule nadbubrežne žlijezde.
BIOSINTEZA
Napravljen od aminokiselinatirozin(dolazi sa hranom -dosta u svježem siru, siru, mahunarkama, cokoladi)
Amino kiseline fenilalanin(radi isto) Fenilalanin se pretvara u tirozinu jetri.
FA hidroksilaza T hidroksilaza DOPA dekarboksilaza
Fenilalanin - Tirozin - Dioksifenilalanin - Dopamin
(DOPA) DOPAMIN hidroksilazaNorepinefrin
METILtransferaza
Adrenalin
Na dopaminergičkim sinapsamabiosinteza medijatora ide do dopamina.
Na noradrenergičkim sinapsama -na norepinefrin (već u granulama).
Na adrenergičkim sinapsama -na adrenalin (neurone nekih područja centralnog nervnog sistema,
srž nadbubrežne žlijezde).
DEPOZIT
Taloženje kateholaminau granulamanastaje zbog vezivanja za određeni protein i ATP. Postojitri bazenakateholamini u nervnim završecima.Rezervni bazen: u granulama, koji se ne oslobađaju po prijemu nervnog impulsa
dok se preostali bazeni ne iscrpe.
Mobilizacija bazen 2 : u granulama, direktno otpuštene
u sinaptičku pukotinu po prijemu impulsa
Mobilizacija bazen 1 : istrošeni transmiter reapsorbovan iz sinaptike
pukotine i višak transmitera zbog zasićenja granula.
Između tri bazena postoji dinamička ravnoteža.
PUSTITE U SYNAPTIC CLEAN
INTERAKCIJA SA RECEPTOROM
receptor:Ahlquist unutra 1948. On je sugerirao da kateholamini djeluju na nekoliko tipova receptora.
Sad:a1, a2, b1, b2, b3 podtipovi
Lokalizacija:
postsinaptička membrana,
presinaptička membrana,
Vanjske sinapse (u organima koji ne primaju presinaptičku inervaciju)
REVERSE CAPTURE
Recapture izloženi 80% posrednik
(nedostatak supstrata, energetski intenzitet sinteze medijatora)
INAKTIVACIJA POSREDNIKA
Inaktivacija izloženi 20%.
Inaktivacija : 1) Oksidativna deaminacija preko mitohondrijalnog enzima MAO - 5%
u sinaptičkom pukotinu.
2) Metilacija sa enzim COMT - 15% ,
koji je ugrađen u postsinaptičke membrane.
ADRENERGIČNI LIJEKOVI
DIRECT ACTION
Djelujte direktno na adrenergičkih receptora .INDIREKTNA AKCIJA
Simpatolitici i simpatomimeticiAfekt osloboditi ili deponovati posrednika.
ADRENORESEPTORI
Alfa adrenergički receptori
Lokalizacija | Efekti aktivacije |
| 1 | |
| Sudovi kože, sluzokože, unutrašnji organi (prekapilarne arteriole), krvni sudovi | Spazam, povećan periferni vaskularni otpor i krvni pritisak |
| Radijalni mišić šarenice | Midriaz |
| Glatki mišići crijeva | Opuštanje |
| Sfinkteri gastrointestinalnog i genitourinarnog trakta | Spazm |
| Miometrijum | Spazm |
| Glatki mišići prostate | Spazm |
| Jetra | Aktivacija glikogenolize |
| Motori pile | Piloerekcija |
| 2 | |
| Terminali adrenergičkih i holinergičkih neurona (presinaptički receptori u centralnom nervnom sistemu i na periferiji) | Smanjenje oslobađanja neurotransmitera (noradrenalin i drugi) |
Presynaptic | |
Vasomotorni centaroblongata medulla | Smanjena aktivnost vazomotornog centra, smanjenje krvnog pritiska |
| Postsynaptic | |
| Žile kože, sluzokože | Spazm |
| Pokretljivost i tonus gastrointestinalnog trakta i crijeva | Smanjenje |
| Extrasynaptic receptori u krvnim sudovima | Vazokonstrikcija |
| Beta ćelije pankreasa | Smanjeno lučenje insulina |
| Trombociti | Agregacija trombocita |
Beta adrenergički receptori
Lokalizacija | Efekti aktivacije |
|
| 1 | ||
| Srce | Tahikardija, povećan minutni volumen i brzina AV provođenja | |
| Jukstaglomerularne ćelije bubrega | Povećano lučenje renina | |
| CNS | Aktivacija vazomotornog centra | |
| Masno tkivo | Aktivacija lipolize | |
| 2 | ||
| Bronhi | Bronhijalna dilatacija |
|
| Žile skeletnih mišića | Ekspanzija, smanjenje krvnog pritiska |
|
| Miometrijum | Opuštenost, smanjena razdražljivost |
|
| Jetra | Aktivacija glikogenolize |
|
| Pankreas β-ćelije Langerhansovih otočića | Oslobađanje insulina |
|
| 3
| ||
| Masno tkivo | Aktivacija lipolize |
|
ADR E N O M I M E T I K
a-ADRENOMIMETIKA
α 1 - adrenergički agonisti
Efekti
-krvni sudoviŽile kože i sluzokože (u većoj mjeri)
Abdominalni organi
Skeletni mišić
Mozak i srce (manje, jer njima dominirajuu 2- vazodilatatorski receptori)
Mezaton
Nije kateholamin (sadrži samo 1 hidroksilnu grupu u aromatičnom prstenu). Niska izloženost COMT - više dugo Efekat. Preovlađuje dejstvo na krvne sudove.
Efekti
1.Suženje krvnih sudova.2. Dilatacija zenice (aktivira a1 receptore radijalni mišići šarenice)
3. Smanjenje intraokularnog pritiska (Povećava odliv intraokularne tečnosti).
Aplikacija
1. Liječenje akutne hipotenzije 0,1-0,5 ml 1% rastvora u 40 ml 5-40% rastvora glukoze2. Rinitis, konjuktivitis. 0,25% -0,5% rastvor
3. S lokalnim anesteticima(za smanjenje resorptivnog efekta)
4. Pregled fundusa
proširenje zjenica (manje trajnije od atropina)
5. Liječenje glaukoma otvorenog ugla.
α 2 - adrenergički agonisti
Mehanizam djelovanja
Stimulacija presinaptičkih α 2 -adrenergičkih receptora u centralnom nervnom sistemu (inhibicija).Ovi receptori, stabilizirajući presinaptičku membranu, smanjuju oslobađanje transmitera
(noradrenalin, dopamin i ekscitatorne aminokiseline - glutaminska, asparaginska).
Hipotenzivni efekat zahvaljujući smanjeno oslobađanje norepinefrina u presorske neurone SDC.
Ovo smanjuje centralni simpatički tonus i povećava tonus vagusa.
Lokalizacija α2 receptora i efekti njihove stimulacije
Medulla– smanjen tonus simpatičkog nervnog sistema, povećan tonus vagusnog nerva.Cerebralni korteks– sedacija, pospanost.
Trombociti- agregacija
Pankreas– inhibicija lučenja insulina.
presinaptička membrana- smanjuju oslobađanje norepinefrina iz završetaka simpatičkih nerava. Povećano oslobađanje acetilholina iz parasimpatičkih nervnih završetaka.
Nuspojave agonista α2 receptora
Posljednjih godina ovi lijekovi se rijetko koriste, zbog njihove loše podnošljivosti.
Suva usta
Sedacija (pospanost, opća slabost, oštećenje pamćenja),
depresija,
Nazalna kongestija,
ortostatska hipotenzija,
Zadržavanje tečnosti
Seksualna disfunkcija.
klonidin (a 2)
Glavni efekti :1.Antihipertenziv . Uvjetuje:
1) inhibicija presorskog dijela vazomotornog centra
2) smanjeno lučenje kateholamina nadbubrežne žlezde
3) privremeni smanjena proizvodnja renina
Posebnost –
kratkotrajno povećanje krvnog tlaka uz brzu intravensku primjenu
zbog stimulacije ekstrasinaptičkih alfa-2 adrenergičkih receptora krvnih sudova
(čak i prije nego što lijek uđe u centralni nervni sistem).
Traje 5-10 minuta.
Potrebne su individualne doze i režimi.
2.Smanjen intraokularni pritisak.
Koristi se za glaukom otvorenog ugla - kapi.
3.Analgetski efekat.
Zbog aktivacije α2-adrenergičkih receptora C i Aδ vlakana
dorzalnih rogova kičmene moždine i moždanog stabla.
Povećava oslobađanje enkefalina i β-endorfina.
Nuspojave
Tolerancija razvija se nakon nekoliko sedmica kontinuirane upotrebe.
Sindrom povlačenja
Naglo povlačenje klonidina dovodi do oslobađanja norepinefrina,deponovan u adrenergičkim završecima.
Ovo je popraćeno
Psihoemocionalno uzbuđenje,
arterijska hipertenzija,
tahikardija,
aritmija,
Substernalna i glavobolja.
18-36 sati nakon posljednje doze, traje 1-5 dana
Prevencija sindroma ustezanja- postepeno smanjenje doze (najmanje 7 dana),
bolje pod maskom drugih antihipertenziva.
Izaziva tešku intoksikaciju(toksična doza – 0,004-0,005).
Simptomi intoksikacije:
Letargija, teška slabost,
hipotermija,
Glavobolja,
Hipotonija skeletnih mišića, hiporefleksija,
Suženje zjenica,
Suhe sluzokože,
Respiratorna depresija
ortostatska hipotenzija,
Bradikardija, atrioventrikularni blok, koma.
Aplikacija :
Ublažavanje hipertenzivne krize
Sublingvalno, sporo intravenozno (rijetko), flaster.
naftizin, galazolin (a 2)
Vazokonstriktorski efekat je snažan i dugotrajan.
Aplikacija
Anti-edematozno, protuupalno dejstvo –
za olakšavanje nazalnog disanja tokom rinitis, za zaustavljanje krvarenja iz nosa.
β-ADRENOMIMETIKA
dobutamin ( V 1
)
Mehanizam djelovanja
Aktivira u 1-adrenoreceptorisrca(Povećava kontraktilnost miokarda i minutni volumen srca).Tahikardija je slabo izražena - zbog refleksne aktivacije vagalnih utjecaja na sinusni čvor.
(iz baroreceptora luka aorte)
Nema značajnog povećanja krvnog pritiska (zbog blage aktivacijeu 2- receptori.
Aplikacija
Akutno zatajenje srca (slabljenje kontraktilne funkcije miokarda).Fenoterol=Berotec=Partusisten ( V 2 )
Selektivnije akcija na u 2 -adrenoreceptori.
Aplikacija
Bronhodilatator. Aerosol, tablete, sirup.Jače i dugotrajnije dejstvo kod bronhospastičnih stanja.
0,1% rastvor za inhalaciju u bocama od 20 ml (0,5 ml po inhalaciji)
Partusisten
U akušerskoj praksi (opušta se mišići materice).
Orciprenaline=Alupent ( v1, v2)
Relativno selektivno akcija na V 2 - bronhijalni receptori.
Aplikacija
Za ublažavanje napada bronhijalne astme može se primijeniti 1-2 ml 0,05% otopine intramuskularno i subkutano.Nakon inhalacije, efekat je u roku od 10-15 minuta, maksimalno sat vremena i do 4-5 sati.
Izadrin ( v1, v2)
Aktivira u 1 srca i u 2 - bronhijalne adrenergičke receptore.
Izražena stimulacija rada srca(tahikardija, intenziviranje
metabolički procesi,
značajan povećana potreba miokarda za kiseonikom,
ali i poboljšana isporuka O2 zbog proširenja koronarnih sudova).
Može se brzo razviti iscrpljenost funkcionalne i metaboličke rezerve srca.
Stimuliše provodni sistem srca - povećana ekscitabilnost i automatizam (aritmije).
Proširuje periferne krvne sudove,snižavanje krvnog pritiska.
Najaktivniji bronhodilatator
od poznatih droga.
a, c – ADrenomimetici
adrenalin ( A 1
A 2
V 1
,V 2
)
norepinefrin ( A 1 A 2 V 1 )
Uticaj na srce
Uticaj nau 1- receptori provodnog sistema.
Oni pobuđuju sinusni čvor srca (manje norepinefrina), povećavaju automatizam.
Broj otkucaja srca se povećava.
Adrenalin
u slučaju srčanog zastoja, ubrizgava se u šupljinu lijeve komore
u kombinaciji sa masažom srca (tako da adrenalin sa krvlju uđe u koronarne arterije i dođe do sinusnog čvora).
Povećava se tonus miokarda.
Minutni volumen i rad srca se povećavaju.
Potrošnja kiseonika miokarda naglo raste.
Efikasnost srca (rad/potrošnja O2) se smanjuje
Može doći do iscrpljivanja srčanih rezervi i razvoja akutnog zatajenja srca.
Utjecaj na krvne sudove
Kontrakcija perifernih sudova, zatim velikih vena i arterija.
Kao rezultat, povećava se povratak krvi u srce.
Žile plućne cirkulacije manje reagiraju, ali čak i u njima
pritisak raste (može se razviti adrenalinski plućni edem).
U žilama skeletnih mišića oni prevladavajuu 2-receptori - vazodilatacijski efekat adrenalina. (Ukupni vaskularni kapacitet skeletnih mišića je veliki -dijastolni pritisak obično se smanjuje).
Sistolni pritisak krv se povećava zbog naglog povećanja funkcije srca.
Norepinefrinza razliku od adrenalina
povećava krvni pritisak uglavnom zbog vazokonstrikcije.
Pogodniji za liječenje akutne hipotenzije.
Utjecaj na tonus glatkih mišića bronha.
Adrenalin (noradrenalin slab)
smanjuje akutno oticanje sluznice.
Koristi se ako su druga sredstva neefikasna. Bolje - inhalacija.
Utjecaj na metabolizam ugljikohidrata.
adrenalin -inzulinski antagonist.
Dramatično povećava razgradnju glikogena u glukozu.
Norepinefrinpraktično nema efekta.
Penetracija kroz BBB
I jedno i drugo slabo prodire.Validan manje od 10 minuta.
SYMPATOMIMETICS
Efedrin hidrohlorid
Pozivi oslobađanje norepinefrina iz presinaptičkih terminala,kao rezultat, sve vrste adrenergičkih receptora su indirektno stimulirane.
U poređenju sa adrenalinom
Manje aktivira alfa adrenergičke receptore
Shodno tome, manje povećava krvni pritisak.
Dobro prodire kroz BBB.
Može izazvati ovisnost i ovisnost.
Aplikacija :
Ublažavanje i prevencija napada astme kod svih vrsta bronhijalne astme.
Rijetko se koristi samostalno zbog nuspojava.
Uključeno u različite kombinovane lekove: Teofedrin, Solutan, Bronholitin.
Nuspojave
Izaziva vazokonstrikciju, povišen krvni pritisak, proširenje bronha, proširenje zjenice, inhibiciju crijevne pokretljivosti.Renders specifično stimulativno dejstvo na centralni nervni sistem
(euforija).
Efikasan kada se uzima oralno.
Kokain
Aplikacija ograničena - lokalna anestezija konjunktive, rožnice
Izaziva vazokonstrikciju u području primjene.
Ima izraženo dejstvo na centralni nervni sistem (euforija)
Tolerancija brzo se razvija, zavisnik može uzimati veće doze u odnosu na terapeutske.
INDIKACIJE ZA UPOTREBU ADrenomimetika
1. Hipotenzija različitog porekla.Norepinefrin, dopamin, mezaton.
Akutno zatajenje srca.Dobutamin.
Otkazivanje Srca.Adrenalin.
Atrioventrikularni blok.Isadrin, orciprenalin.
Bronhijalna astma.Salbutamol, fenoterol, orciprenalin, efedrin.
Rizik od pobačaja.Partusisten = Fenoterol.
Neki oblici glaukoma (otvoreni ugao)Mezaton, klonidin, adrenalin.
Da produži efekat MA.Adrenalin, mesaton.
Hitno liječenje anafilaktičkog šoka.Adrenalin.
Hipoglikemijska koma.Adrenalin.
NUSPOJAVE
a- Adrenergički agonisti
Opasan porast krvnog pritiska. Posljedica je oštro preopterećenje srca, njegova iscrpljenost,
akutno zatajenje srca s razvojem plućnog edema.
c- Adrenergički agonisti
Poremećaji srčanog ritma, napad angine, tremor mišića.
ADRENOLITICI I SIMPATOLITICI
Adrenolitici blok adrenoreceptori.
Eliminirati ili spriječiti efekte adrenergičkih agonista.
Simpatoliticičin na presinaptičkom nivou
.
Smanjuje otpuštanje medijatora.
(promjena njihove sinteze, skladištenja i oslobađanja).
Ne blokiraju adrenergičke receptore.
Ne eliminirati efekte kateholamina koji se primjenjuju izvana.
SYMPATHOLYTICS
Konačan rezultatefekti simpatolitika - slabljenje prijenosa impulsa
od završetaka simpatičkih nerava do odgovarajućih organa.
Zahvaljujući .
smetnje u sintezi medijatora
. nedostatak norepinefrina
. blokada oslobađanja neurotransmitera
Kao rezultat . vaskularni tonus se smanjuje
. smanjuju se refleksne reakcije kardiovaskularnog sistema
za razne podsticaje
. krvni pritisak se smanjuje
. smanjenje metaboličkih promjena,
Adrenoreceptori organa (sudovi, srce)
potpuno zadržavaju osjetljivost na kateholamine
Najvažniji Efekatsimpatolitici -antihipertenziv.
Metildopa
Mehanizam djelovanja
1. Kompetitivni je biohemijski antagonist DOPA (dioksifenilalanin) –
prekursor dopamina i norepinefrina i odlaže njihovu sintezu.
U tijelu se prvo pretvara u metildopamin, a zatim u metilnorepinefrin,
formiranje “lažnih” manje aktivnih medijatora.
2.Metilnorepinefrin je selektivni alfa-2 adrenergički agonist –
ovo objašnjava centralnu komponentu antihipertenzivnog efekta.
Konačan efekat- aktivacija “negativne povratne sprege” u regulaciji oslobađanja NA i smanjenje centralnog simpatičkog vaskularnog tonusa
Glavni efekti
Antihipertenzivni efekat zahvaljujući
Vazodilatacija i smanjenje OPS
Nuspojave
Pored toga, nuspojave agonista α2 receptora
Može ometati dopaminergičke mehanizme supresije lučenja prolaktina
(sekrecija se povećava)
stoga, kada se koriste u nekim slučajevima, muškarci razvijaju ginekomastiju,
a kod žena - galaktoreja.
Sindrom povlačenja, moguća pojava poremećaja srčanog ritma.
Disfunkcija jetre.
Hemolitička anemija.
Aplikacija
Liječenje hipertenzije.
Rezerpin
Mehanizam djelovanja
1. Odgađa apsorpciju granula prekursora norepinefrina - dopamina,koje oksidira MAO.
Blokira povratak "potrošenog" noradrenalina u granule,
Posljedica: Fond kateholamina u granulama je iscrpljen.
Efekti
1.Polagano se razvija umjereno hipotenzivno djelovanje.
Traje 1-3 mjeseca nakon prestanka uzimanja lijeka.
2. Psihosedativni efekat.
U dozama 2-3 puta većim od antihipertenzivnih, rezerpin ublažava manifestacije patologije na nivou psihoze.
Zasnovan je na sposobnosti blokiranja aktivirajućeg utjecaja na više dijelove mozga noradrenergičkih, dopaminergičkih uzlaznih aksona iz neurona matičnih struktura.
3. Vagotonična akcija.
Rezultat blokiranja simpatičkog prijenosa na periferiji i povećanja reaktivnosti vagalnih centara.
To se manifestuje u vidu bradikardije, povećanog tonusa i sekrecije želuca, motiliteta crijeva i bronhijalnog tonusa.
Octadine
Mehanizam djelovanja
1. Inhibicija aktivnog povratka kateholamina iz sinaptičkog pukotina,Kao rezultat toga, COMT ih deaktivira.
2. Sposobnost taloženja u citosolu i granulama adrenergičkih završetaka,
ističući se kao neaktivni “lažni posrednici”.
To dovodi do iscrpljivanja medijatorskog fonda uz njegov spor oporavak nakon povlačenja.
Efekti
Smanjenje krvnog pritiska (dilatacija kapacitivnih žila i slabljenje srčanih reakcija).Nuspojave
Do kolapsa dolazi lako kada se promijeni položaj tijela.Ornid
Mehanizam djelovanja
1. Blokada kalcijumskih kanala presinaptičke membrane i spojna funkcija kalcijuma u mehanizmu oslobađanja transmitera iz granula.
Kao rezultat toga, čini se da ornid "zaključava" neurotransmiter u simpatičnom završetku
manifestuje se: razvojem hipertenzivne krize
napadi angine,
napadi aritmije.
Povećani nivoi aterogenih lipida u krvi.
Seksualna disfunkcija kod muškaraca
učestalost od 11 do 28% uz dugotrajnu upotrebu propranolola uzavisno od doze
Neželjeni efekti na centralninervni sistem:
nesanica, noćne more, halucinacije, mentalna depresija.
Aplikacija
Terapija hipertenzije.
IHD terapija
Terapija aritmije
SPISAK LIJEKOVA NA RECEPT
Zbog izuzetnog značaja adrenergičkog nervnog sistema za stimulaciju kontraktilnosti zdravog miokarda, proučavana je njegova aktivnost kod pacijenata sa kongestivnom srčanom insuficijencijom. Aktivnost ovog sistema u mirovanju i tokom fizičke aktivnosti procenjivana je koncentracijom norepinefrina u arterijskoj krvi. Kod zdravih osoba, postoji relativno mali porast nivoa norepinefrina tokom vežbanja. Kod pacijenata sa srčanom insuficijencijom, cirkulirajući nivoi norepinefrina, čak i u mirovanju, mogu biti značajno povišeni. Štoviše, prognoza bolesti je lošija što je veća koncentracija neurotransmitera. Osim toga, kod pacijenata sa kongestivnom srčanom insuficijencijom, tokom fizičke aktivnosti, sadržaj norepinefrina u krvi raste u znatno većoj mjeri nego kod zdravih ljudi. Ovo se objašnjava i značajno većom aktivnošću adrenergičnog nervnog sistema kod ove grupe pacijenata, koja se zadržava i tokom fizičke aktivnosti.
Važnost povećanja aktivnosti adrenergičkog nervnog sistema za održavanje ventrikularne kontraktilnosti u uslovima suprimirane funkcije miokarda kod kongestivne srčane insuficijencije potkrepljuju podaci da blokada beta-adrenergičkih receptora može pogoršati disfunkciju pumpe. Stoga, adrenergični nervni sistem igra važnu modulirajuću ulogu u održavanju cirkulacije krvi kod pacijenata sa kongestivnom srčanom insuficijencijom. S tim u vezi, antiadrenergičke lijekove, posebno beta-blokatore, treba koristiti s velikim oprezom pri liječenju pacijenata sa ograničenom rezervom miokarda (Poglavlje 182).
Na istom. s vremenom se koncentracija i sadržaj norepinefrina u tkivima srca kod pacijenata sa srčanom insuficijencijom smanjuje i u nekim slučajevima iznosi samo 10% normalnih vrijednosti. Mehanizam koji leži u osnovi ovog fenomena je potpuno nepoznat. Međutim, vjeruje se da dugotrajno očuvanje visokog tonusa simpatičkih nerava srca igra odlučujuću ulogu, ometajući na određeni način biosintezu norepinefrina. Osim toga, postoje dokazi da se kod hronične teške srčane insuficijencije značajno smanjuju gustina b-adrenergičkih receptora u srcu i koncentracija cikličkog AMP u miokardu.
S obzirom na snažan pozitivan inotropni efekat norepinefrina koji oslobađaju ovi nervi, adrenergični nervni sistem se može smatrati važnim potencijalnim izvorom za održavanje funkcije otpalog miokarda. Međutim, povećanje učestalosti i jačine srčanih kontrakcija kod životinja s eksperimentalnim zatajenjem srca i iscrpljivanjem rezervi norepinefrina u srcu praktički izostaje ili je minimalno izražen kada su stimulirani simpatički nervi srca. Dakle, čini se da je u slučajevima kada je kongestivno zatajenje srca praćeno iscrpljivanjem rezervi norepinefrina u srcu, količina iste koju oslobađaju simpatički nervni završeci u srcu mala u odnosu na impulse koji se prenose ovim nervima. Štaviše, čak ni oslobođeni norepinefrin ne može imati odgovarajući efekat na miokard zbog inhibicije efektivnog adrenergičkog mehanizma miokarda.
Istovremeno, prisustvo rezervi norepinefrina u miokardu nije preduslov za održavanje njegove kontraktilnosti. Međutim, budući da je smanjenje zaliha norepinefrina u miokardu kod zatajenja srca kombinovano sa smanjenjem oslobađanja ovog neurotransmitera, može se pretpostaviti da ovo smanjenje potonjeg leži u osnovi gubitka tako neophodne adrenergičke podrške za oštećenu funkciju miokarda. U kasnijim fazama srčane insuficijencije, kada su razine kateholamina u cirkulaciji povišene i nivoi norepinefrina u miokardu smanjeni, miokard postaje u velikoj mjeri ovisan o generaliziranoj adrenergičkoj stimulaciji iz ekstrakardijalnih izvora, uglavnom nadbubrežne moždine. Ova činjenica objašnjava pogoršanje srčane funkcije koje se javlja kod pacijenata sa srčanom insuficijencijom koji primaju beta blokatore. Ova generalizirana adrenergička stimulacija, koja je rezultat cirkulacije velikih količina kateholamina u krvi, može, međutim, imati štetne nuspojave povezane s povećanjem vaskularnog otpora, a time i naknadnog opterećenja, koje značajno premašuje optimalne vrijednosti.
Završavajući analizu mehanizama srčane insuficijencije, treba napomenuti da glavni poremećaji leže u potiskivanju odnosa između snage i brzine srčanih kontrakcija i pomaka krivulje dužine aktivne napetosti srčanih vlakana. Ovo odražava smanjenje kontraktilnosti miokarda (vidi sliku 181-6, krive 1, 3). U mnogim slučajevima, minutni volumen srca i vanjski rad ventrikula kod ovih pacijenata u mirovanju ostaju u granicama normale, što se, međutim, osigurava samo povećanjem krajnje dijastoličke dužine mišićnih vlakana i povećanjem krajnjeg dijastoličkog volumena ventrikula, tj. zbog mehanizma Frank - Starling (vidi sl. 181-6, tačke A-D). Povećanje predopterećenja lijeve komore praćeno je sličnim promjenama plućnog kapilarnog tlaka, što uzrokuje kratak dah kod pacijenata sa srčanom insuficijencijom. Kontraktilnost miokarda zbog povećane simpatičke aktivnosti tokom fizičke aktivnosti kod pacijenata sa teškom srčanom insuficijencijom se ne povećava ili se povećava u maloj meri, što je posledica iscrpljivanja rezervi norepinefrina u miokardu (vidi sliku 181-6, krive 3 i 3). Mehanizmi koji održavaju punjenje ventrikula krvlju tokom fizičke aktivnosti kod zdravih osoba dovode do daljeg pogoršanja funkcije miokarda kada je ona nedovoljna, što rezultira izravnavanjem krivulje napetosti vlakana aktivne dužine. I unatoč činjenici da lijeva komora nakon njihovog uključivanja može malo poboljšati svoju aktivnost, ovaj učinak se postiže isključivo zbog pretjeranog povećanja krajnjeg dijastoličkog volumena i tlaka lijeve klijetke, a time i tlaka u plućnim kapilarama. Potonji faktor dovodi do pojačanog nedostatka zraka, što zauzvrat igra važnu ulogu u ograničavanju intenziteta fizičke aktivnosti koju obavlja pacijent. Zatajenje lijeve komore postaje ireverzibilno kada je kriva dužine aktivne napetosti mišićnih vlakana toliko depresivna da srčana aktivnost nije u stanju da zadovolji metaboličke potrebe perifernih tkiva u mirovanju (vidi sliku 181-6, krivulja 4) i/ili na kraju -dijastolni pritisak u levoj komori i pritisak u plućnim kapilarama raste do te mere da dovodi do razvoja plućnog edema (vidi sliku 181-6, tačka E).
Posljednjih godina veliki broj radova posvećen je kateholaminima i srodnim spojevima. To je posebno zbog činjenice da su interakcije između endogenih kateholamina i niza lijekova koji se koriste u liječenju hipertenzije, mentalnih poremećaja i dr. izuzetno važne za kliničku praksu.O ovim lijekovima i interakcijama će se detaljnije govoriti u nastavku. poglavlja. Ovdje ćemo analizirati fiziologiju, biohemiju i farmakologiju adrenergičkog prijenosa.
Sinteza, skladištenje, oslobađanje i inaktivacija kateholamina
Slika 6.3. Sinteza kateholamina.
Sinteza. Pretpostavku o sintezi adrenalina iz tirozina i redoslijedu faza ove sinteze (slika 6.3) prvi je iznio Blashko 1939. godine. Od tada su svi odgovarajući enzimi identificirani, karakterizirani i klonirani (Nagatsu, 1991). Važno je da svi ovi enzimi nemaju apsolutnu specifičnost pa stoga i druge endogene supstance i lijekovi mogu sudjelovati u reakcijama koje katalizuju. Dakle, dekarboksilaza aromatične L-amino kiseline (DOPA dekarboksilaza) može katalizirati ne samo konverziju DOPA u dopamin, već i 5-hidroksitriptofan u serotonin (5-hidroksitriptamin) i metildopa u α-metildopamin; potonji se pod utjecajem dopamin-β-monooksigenaze (dopamin-β-hidroksilaze) pretvara u "lažni posrednik" - a-metilnorepinefrin.
Smatra se da je ograničavajuća reakcija u sintezi kateholamina hidroksilacija tirozina (Zigmond et al., 1989). Enzim tirozin hidroksilaza (tirozin-3-monooksigenaza) koji katalizuje ovu reakciju aktivira se stimulacijom adrenergičkih neurona ili ćelija medule nadbubrežne žlijezde. Ovaj enzim služi kao supstrat protein kinaze A (cAMP-zavisne), Ca2+-kalmodulin-zavisne protein kinaze i protein kinaze C. Smatra se da upravo njegova fosforilacija pod uticajem protein kinaza dovodi do povećanja njegove aktivnosti. (Zigmond et al., 1989; Daubner et al., 1992). Ovo je važan mehanizam za povećanje sinteze kateholamina uz pojačanu aktivnost simpatičkih nerava. Uz to, stimulacija ovih nerava je praćena odgođenim povećanjem ekspresije gena tirozin hidroksilaze. Postoje dokazi da ovo povećanje može biti posljedica promjena na različitim nivoima - transkripcije, obrade RNK, regulacije stabilnosti RNK, translacije i stabilnosti samog enzima (Kumer i Vrana, 1996). Biološko značenje ovih efekata je da se povećanim oslobađanjem kateholamina održava njihov nivo u nervnim završecima (ili ćelijama medule nadbubrežne žlezde). Osim toga, aktivnost tirozin hidroksilaze može biti inhibirana kateholaminima kroz mehanizam alosterične modifikacije; stoga, ovdje postoje negativne povratne informacije. Mutacije gena tirozin hidroksilaze su opisane kod ljudi (Wevers et al., 1999).
Opis za sl. 6.3. Sinteza kateholamina. Desno od strelica su enzimi (kurzivom) i kofaktori. Posljednja faza (formiranje adrenalina) događa se samo u meduli nadbubrežne žlijezde i nekim neuronima koji sadrže adrenalin u moždanom stablu.
Naša saznanja o mehanizmima i lokalizaciji u ćeliji procesa sinteze, skladištenja i oslobađanja kateholamina zasnivaju se na proučavanju organa sa simpatičkom inervacijom i medule nadbubrežne žlijezde. Što se tiče organa sa simpatičkom inervacijom, gotovo sav norepinefrin koji se nalazi u njima lokaliziran je u nervnim vlaknima - nekoliko dana nakon transekcije simpatičkih živaca, njegove rezerve su potpuno iscrpljene. U ćelijama medule nadbubrežne žlijezde kateholamini se nalaze u takozvanim hromafinskim granulama (Winkler, 1997; Aunis, 1998). To su vezikule koje sadrže ne samo kateholamine u izuzetno visokim koncentracijama (oko 21% suhe težine), već i ATP i niz proteina – hromogranine, dopamin-β-monooksigenazu, enkefaline, neuropeptid Y i druge. Zanimljivo je da N-terminalni fragment hromogranina A, vazostatin-1, ima antibakterijska i antifungalna svojstva (Lugardon et al., 2000). Na završecima simpatičkih nerava pronađene su dvije vrste vezikula: velike vezikule guste elektronima, koje odgovaraju hromafinskim granulama, i male mjehuriće guste elektronima koje sadrže norepinefrin, ATP i dopamin-β-monooksigenazu vezanu za membranu.

Slika 6.4 Osnovni mehanizmi sinteze, skladištenja, oslobađanja i inaktivacije kateholamina.
Glavni mehanizmi sinteze, skladištenja, oslobađanja i inaktivacije kateholamina prikazani su na Sl. 6.4. U adrenergičnim neuronima, enzimi odgovorni za sintezu norepinefrina se formiraju u tijelu i transportuju duž aksona do završetaka. U citoplazmi se dešavaju hidroksilacija tirozina u DOPA i dekarboksilacija DOPA u dopamin (slika 6.3). Zatim se približno polovina nastalog dopamina aktivnim transportom prenosi u vezikule koje sadrže dopamin-β-monooksigenazu, i tu se dopamin pretvara u norepinefrin. Preostali dopamin prvo se podvrgava deaminaciji (da nastane 3,4-dihidroksifeniloctena kiselina), a zatim O-metilaciji (da nastane homovanilna kiselina). U meduli nadbubrežne žlijezde postoje 2 tipa ćelija koje sadrže kateholamine: norepinefrin i adrenalin. Potonji sadrže enzim feniletanolamin-N-metiltransferazu. U ovim ćelijama norepinefrin ostavlja hromafinske granule u citoplazmu (očigledno difuzijom) i tu se metilira navedenim enzimom u adrenalin. Potonji ponovo ulazi u granule i čuva se u njima do oslobađanja. Kod odraslih, adrenalin čini oko 80% svih kateholamina u srži nadbubrežne žlijezde; preostalih 20% je pretežno norepinefrin (von Euler, 1972).
Opis za sl. 6.4. Osnovni mehanizmi sinteze, skladištenja, oslobađanja i inaktivacije kateholamina. Prikazan je šematski prikaz simpatičnog završetka. Tirozin se aktivnim transportom prenosi u aksoplazmu (A), gdje se pod djelovanjem citoplazmatskih enzima pretvara u DOPA, a zatim u dopamin (B). Potonji ulazi u vezikule, gdje se pretvara u norepinefrin (B). Akcijski potencijal uzrokuje ulazak u Ca2+ terminal (nije prikazano), što rezultira fuzijom vezikula sa presinaptičkom membranom i oslobađanjem norepinefrina (G). Potonji aktivira α- i β-adrenergičke receptore postsinaptičke ćelije (D) i djelimično ulazi u nju (ekstraneuronski unos); u ovom slučaju, očigledno je inaktiviran konverzijom u normetanefrin pomoću COMT-a. Glavni mehanizam inaktivacije norepinefrina je njegov ponovni unos presinaptičkim terminalom (E) ili neuronski unos. Norepinefrin koji se oslobađa u sinaptički rascjep također može stupiti u interakciju sa presinaptičkim α2-adrenergičkim receptorima (G), potiskujući vlastito oslobađanje (isprekidana linija). Drugi medijatori (na primjer, peptidi i ATP) također mogu biti prisutni na adrenergičkom terminalu - u istim vezikulama kao i norepinefrin, ili u zasebnim vezikulama. AR - adrenoreceptor, DA - dopamin, NA - norepinefrin, NM - normetanefrin, P-peptid
Glavni faktor koji regulira brzinu sinteze adrenalina (a samim tim i sekretornu rezervu medule nadbubrežne žlijezde) proizvodi korteks nadbubrežne žlijezde. Ovi hormoni, preko adrenalnog portalnog sistema, ulaze u visokim koncentracijama direktno u hromafinske ćelije medule i indukuju u njima sintezu feniletanolamin-N-metiltransferaze (slika 6.3). Pod uticajem glukokortikoida povećava se i aktivnost tirozin hidroksilaze i dopamin-β-monooksigenaze u meduli (Carroll et al., 1991; Viskupic et al., 1994). Stoga, dovoljno dugotrajan stres, koji uzrokuje povećanje lučenja ACTH, dovodi do povećanja sinteze hormona kako kortikalnih (uglavnom kortizola) tako i medule nadbubrežne žlijezde.
Ovaj mehanizam djeluje samo kod onih sisara (uključujući ljude) kod kojih su kromafinske stanice medule u potpunosti okružene stanicama korteksa. Kod burbota, na primjer, hromafin i ćelije koje luče steroide nalaze se u zasebnim žlijezdama koje nisu međusobno povezane, a adrenalin se ne luči. Istovremeno, feniletanolamin-N-metiltransferaza kod sisara nalazi se ne samo u nadbubrežnim žlijezdama, već iu nizu drugih organa (mozak, srce, pluća), odnosno moguća je ekstra-nadbubrežna sinteza adrenalina (Kennedy i Ziegler, 1991; Kennedy et al., 1993).
Zalihe norepinefrina u završecima adrenergičkih vlakana obnavljaju se ne samo zbog njegove sinteze, već i zbog ponovnog preuzimanja oslobođenog noradrenalina. U većini organa upravo ponovna pohrana osigurava prestanak djelovanja norepinefrina. U krvnim sudovima i drugim tkivima, gdje su sinaptičke pukotine dovoljno široke, uloga ponovne pohrane norepinefrina nije tako velika – značajan dio se inaktivira ekstraneuronskim unosom (vidi dolje), enzimskim cijepanjem i difuzijom. I ponovni unos norepinefrina u adrenergičke terminale i njegov ulazak u sinaptičke vezikule iz aksoplazme idu suprotno koncentracijskom gradijentu ovog medijatora, te se stoga provode pomoću dva aktivna transportna sistema, uključujući i odgovarajuće nosače. Skladištenje. Budući da su kateholamini pohranjeni u vezikulama, njihovo oslobađanje se može prilično precizno kontrolirati; osim toga, nisu izloženi citoplazmatskim enzimima i ne propuštaju u okolinu. Transportni sistemi biogenih monoamina su prilično dobro proučeni (Schuldiner, 1994). Čini se da je unos kateholamina i ATP-a izolovanim hromafinskim granulama vođen pH i potencijalnim gradijentima koje stvara H+-ATPaza. Prijenos jedne molekule monoamina u vezikulu je praćen oslobađanjem dva protona (Browstein i Hoffman, 1994). Transport monoamina je relativno neselektivni. Na primer, isti sistem je sposoban da transportuje dopamin, norepinefrin, epinefrin, serotonin, kao i meta-1"1-benzilgvanidin, supstancu koja se koristi za izotopsku dijagnostiku tumora iz hromafinskih ćelija feohromocitoma (Schuldiner, 1994). Vezikularni transport amina je potisnut rezerpin, ova supstanca iscrpljuje kateholamine u simpatičkim završecima i mozgu. Metodama molekularnog kloniranja otkriveno je nekoliko cDNK povezanih sa vezikularnim transportnim sistemima.U njima su identifikovani otvoreni okviri za čitanje, što ukazuje na kodiranje proteina sa 12 transmembranskih domena. Ovi proteini moraju biti homologni drugim transportnim proteinima, kao što su oni koji posreduju otpornost bakterija na lijekove (Schuldiner, 1994), a promjene u ekspresiji ovih proteina mogu igrati važnu ulogu u regulaciji sinaptičkog prijenosa (Varoqui i Erickson, 1997).
Kateholamini (na primjer, norepinefrin) koji se unose u krv životinja brzo se akumuliraju u organima s obilnom simpatičkom inervacijom, posebno u srcu i slezeni. U ovom slučaju, označeni kateholamini se nalaze u simpatičkim završecima; desimpatički organi ne akumuliraju kateholamine (vidi pregled Browstein i Hoffman, 1994). Ovi i drugi podaci ukazuju na prisustvo kateholaminskog transportnog sistema u membrani simpatičkih neurona. Utvrđeno je da je ovaj sistem ovisan o Na+ i da ga selektivno blokiraju određeni lijekovi, uključujući kokain i triciklične antidepresive kao što je imipramin. Ima visok afinitet za norepinefrin, a nešto manji za adrenalin. Ovaj sistem ne podnosi sintetički izoprenalin. Neuronsko uzimanje kateholamina naziva se i unos tipa 1 (Iversen, 1975). Metode pročišćavanja proteina i molekularnog kloniranja identificirale su nekoliko visoko specifičnih neurotransmiterskih transportera, posebno transportere visokog afiniteta dopamina, norepinefrina, serotonina i niza aminokiselina (Amara i Kuhar, 1993; Browstein i Hoffman, 1994; Masson et al. , 1999). Svi oni pripadaju velikoj porodici proteina, čije zajedničke karakteristike uključuju, na primjer, 12 transmembranskih domena. Očigledno je specifičnost membranskih transportera veća od one vezikularnih. Osim toga, ovi transporteri služe kao mjesta primjene za supstance kao što su (transporter dopamina) i (transporter dopamina).
Takozvani indirektni simpatomimetici (na primjer, tiramin) ispoljavaju svoje djelovanje indirektno, obično izazivajući oslobađanje norepinefrina iz simpatičkih završetaka. Dakle, aktivni princip u propisivanju ovih lijekova je sam norepinefrin. Mehanizmi djelovanja indirektnih simpatomimetika su složeni. Svi se vezuju za transportere koji osiguravaju neuronsko preuzimanje kateholamina i zajedno s njima prelaze u aksoplazmu; u ovom slučaju, nosač se pomiče na unutrašnju površinu membrane i na taj način postaje dostupan za norepinefrin (metabolički olakšana difuzija). Osim toga, ovi lijekovi uzrokuju oslobađanje norepinefrina iz vezikula, natječući se s njim u vezikularnom transportnom sistemu. Rezerpin, koji uzrokuje iscrpljivanje norepinefrina u vezikulama, također blokira vezikularni transport, ali, za razliku od indirektnih simpatomimetika, ulazi u terminal jednostavnom difuzijom (Bonish i Trendelenburg, 1988).
Prilikom propisivanja indirektnih simpatomimetika često se opaža ovisnost (tahifilaksa, desenzibilizacija). Stoga, kada se tiramin uzima više puta, njegova efikasnost opada prilično brzo. Nasuprot tome, ponovljena primjena norepinefrina nije praćena smanjenjem djelotvornosti. Osim toga, eliminira se ovisnost o tiraminu. Ne postoji definitivno objašnjenje za ove pojave, iako su iznesene neke hipoteze. Jedna od njih je da je dio norepinefrina koji je istisnut indirektnim simpatomimetima mali u odnosu na ukupne rezerve ovog medijatora u adrenergičkim završecima. Pretpostavlja se da ova frakcija odgovara vezikulama koje se nalaze uz membranu, a iz njih je norepinefrin istisnut manje aktivnim indirektnim simpatomimetikom. Kako god bilo, indirektni simpatomimetici ne izazivaju oslobađanje iz terminala dopamin-β-monooksigenaze i mogu djelovati u okruženju bez kalcija - što znači da njihov učinak nije povezan s egzocitozom.
Postoji i sistem za ekstraneuronski unos kateholamina (uptake tip 2), koji ima nizak afinitet za norepinefrin, nešto veći afinitet za adrenalin i još veći afinitet za izoprenalin. Ovaj sistem je sveprisutan: nalazi se u ćelijama glija, jetre, miokarda i drugih ćelija. Imipramin i kokain ne blokiraju ekstraneuronsko preuzimanje. U uslovima neometanog neuronskog preuzimanja, njegova uloga je naizgled mala (Iversen, 1975; Trendelenburg, 1980). Možda je važnije za uklanjanje kateholamina iz krvi nego za inaktivaciju kateholamina koje oslobađaju nervni završeci.
Pustiti. Slijed događaja uslijed kojih se adrenalin oslobađa iz adrenergičkih završetaka pod utjecajem nervnog impulsa nije potpuno jasan. U meduli nadbubrežne žlijezde, pokretački faktor je djelovanje acetilholina koji oslobađaju preganglijska vlakna na N-holinergičke receptore kromafinskih stanica. U tom slučaju dolazi do lokalne depolarizacije, Ca2\ ulazi u ćeliju i eksonitozom se oslobađa sadržaj hromafinskih granula (adrenalin, ATP, neki neuropeptidi i njihovi prekursori, hromogranini, dopamin-β-monooksigenaza). Na adrenergičnim terminalima, ulazak Ca2+ kroz naponsko vođene kalcijumske kanale također igra ključnu ulogu u spajanju depolarizacije presinaptičke membrane (akcioni potencijal) i oslobađanja norepinefrina. Blokada N-tipa kalcijumskih kanala uzrokuje smanjenje AH, očigledno potiskivanjem oslobađanja norepinefrina (Bowersox et al., 1992). Mehanizmi egzocitoze izazvane kalcijumom uključuju visoko konzervirane proteine koji osiguravaju vezivanje vezikula za ćelijsku membranu i njihovu degranulaciju (Aunis, 1998). Povećanje tonusa simpatikusa praćeno je povećanjem koncentracije dopamin-β-monooksigenaze i hromogranina u krvi. Ovo sugerira da je egzocitoza vezikula uključena u oslobađanje norepinefrina nakon stimulacije simpatičkih živaca.
Ako sinteza i ponovna pohrana norepinefrina nije poremećena, onda čak i produžena iritacija simpatičkih nerava ne dovodi do iscrpljivanja rezervi ovog medijatora. Ako se poveća potreba za oslobađanjem norepinefrina, tada u igru stupaju regulatorni mehanizmi. usmjerena, posebno, na aktivaciju tirozin hidroksilaze i dopamin-β-monooksigenaze (vidi gore).
Inaktivacija. Prestanak djelovanja norepinefrina i adrenalina nastaje zbog: 1) ponovnog preuzimanja nervnim završecima, 2) difuzije iz sinaptičkog pukotina i ekstra neuronskog preuzimanja, 3) enzimskog cijepanja. Ovo poslednje izazivaju dva glavna enzima - MAO i COMT (Axelrod, 1966; Kopin, 1972). Osim toga, kateholamini se razgrađuju sulfotransferazama (Dooley, 1998). Istovremeno, uloga enzimske razgradnje u adrenergičkoj sinapsi je mnogo manja nego u holinergičkoj sinapsi, a ponovno preuzimanje zauzima prvo mjesto u inaktivaciji kateholamina. To je vidljivo, na primjer, iz činjenice da blokatori ponovne pohrane kateholamina (kokain, imipramin) značajno pojačavaju efekte norepinefrina, a MAO i COMT inhibitori vrlo slabo. MAO igra ulogu u uništavanju norepinefrina zarobljenog u aksoplazmi. COMT (posebno u jetri) je bitan za inaktivaciju endogenih i egzogenih kateholamina u krvi.
MAOI i COMT su široko rasprostranjeni u cijelom tijelu, uključujući mozak. Njihova koncentracija je najveća u jetri i bubrezima. Istovremeno, COMT je gotovo odsutan u adrenergičkim neuronima. Ova dva enzima se razlikuju i po intracelularnoj lokalizaciji: MAO je pretežno povezan sa vanjskom membranom mitohondrija (uključujući i adrenergičke terminale), a COMT se nalazi u citoplazmi. Svi ovi faktori određuju kojim putem će se kateholamini razgraditi pod različitim uslovima, kao i mehanizmi djelovanja brojnih lijekova. Identifikovana su dva MAO izoenzima (MAO A i MAO B), a njihov odnos u različitim neuronima centralnog nervnog sistema i različitim organima veoma varira. Dostupni su selektivni inhibitori ova dva izoenzima (poglavlje 19). Ireverzibilni inhibitori MAO A povećavaju bioraspoloživost tiramina sadržanog u brojnim namirnicama; Budući da tiramin pojačava oslobađanje norepinefrina iz simpatičkih završetaka, kada se ovi lijekovi kombiniraju s proizvodima koji sadrže tiramin, moguća je hipertenzivna kriza. Selektivni inhibitori MAO B (npr. selegilin) i reverzibilni selektivni inhibitori MAO A (npr. moklobemid) imaju manje šanse da izazovu ovu komplikaciju (Volz i Geiter, 1998; Wouters, 1998). MAO inhibitori se koriste za liječenje Parkinsonove bolesti i depresije (poglavlja 19 i 22).

Slika 6.5. Metabolizam kateholamina. I MAO i COMT su uključeni u inaktivaciju kateholamina, ali redoslijed njihovog djelovanja može biti drugačiji.
Većina epinefrina i norepinefrina koji ulazi u krv, bilo iz nadbubrežne moždine ili adrenergičkih terminala, metilira COMT da bi se formirao metanefrin i normetanefrin, respektivno (slika 6.5). Norepinefrin, oslobođen iz vezikula u aksoplazmu pod uticajem određenih lekova (na primer, rezerpin), prvo se deaminira MAO u 3,4-hidroksimandehid; potonji se reducira aldehid reduktazom u 3,4-dihidroksifeniletilen glikol ili oksidira aldehid dehidrogenazom u 3,4-dihidroksimandelicnu kiselinu. Glavni metabolit kateholamina koji se izlučuje u urinu je 3-metoksi-4-hidroksimandelična kiselina, koja se često (iako netačno) naziva vanililmandelična kiselina. Odgovarajući metabolit dopamina, koji ne sadrži hidroksilnu grupu na bočnom lancu, je homovanilna kiselina. Ostale reakcije metabolizma kateholamina prikazane su na Sl. 6.5. Mjerenje koncentracije kateholamina i njihovih metabolita u krvi i urinu važna je metoda za dijagnosticiranje feohromocitoma (tumor koji luči kateholamine).
MAO inhibitori (npr. pargilin i nialamid) mogu uzrokovati povećane koncentracije norepinefrina, dopamina i serotonina u mozgu i drugim organima, što rezultira različitim fiziološkim efektima. Suzbijanje COMT aktivnosti nije praćeno značajnim reakcijama. Istovremeno, COMT inhibitor entakapon se pokazao prilično efikasnim kod Parkinsonove bolesti (Chong i Mersfelder, 2000; vidi takođe Poglavlje 22).
Opis za sl. 6.5. Metabolizam kateholamina. I MAO i COMT su uključeni u inaktivaciju kateholamina, ali redoslijed njihovog djelovanja može biti drugačiji. U prvom slučaju, metabolizam kateholamina počinje oksidativnom deaminacijom pod utjecajem MAO; Adrenalin i norepinefrin se prvo pretvaraju u 3,4-hidroksimandehid, koji se zatim ili reducira u 3,4-dihidroksifeniletilen glikol ili oksidira u 3,4-dihidroksimandeličnu kiselinu. Prva reakcija drugog puta je njihova metilacija pomoću COMT-a u metanefrin i normetanefrin, respektivno. Zatim djeluje drugi enzim (u prvom slučaju - COMT, u drugom - MAO), a formiraju se glavni metaboliti koji se izlučuju urinom - 3-metoksi-4-hidroksifeniletilen glikol i 3-metoksi-4-hidroksimandelik (vanilil mandelic). ) kiselina. Slobodni 3-met-ci-4-hidroksifeniletilen glikol se u velikoj mjeri pretvara u vanililmandelnu kiselinu. 3,4-dihidroksifeniletilen glikol i, u određenoj mjeri, O-metilirani amini i kateholamini mogu biti konjugirani sa sulfatima ili glukuronidima. Axelrod, 1966, itd.
Klasifikacija adrenergičkih receptora

Tabela 6.3. Adrenergički receptori
Da biste se snašli u neverovatnoj raznolikosti efekata kateholamina i drugih adrenergičkih supstanci, potrebno je dobro poznavati klasifikaciju i svojstva adrenergičkih receptora. Pojašnjenje ovih svojstava i onih biohemijskih i fizioloških procesa na koje utiče aktivacija različitih adrenergičkih receptora pomoglo je da se razumeju različite i ponekad naizgled kontradiktorne reakcije različitih organa na kateholamine. Svi adrenergički receptori su slične strukture (vidi dole), ali su povezani sa različitim sistemima sekundarnih glasnika, pa stoga njihova aktivacija dovodi do različitih fizioloških posledica (tabele 6.3 i 6.4).

Tabela 6.4. Sistemi drugog glasnika povezani sa adrenergičkim receptorima
Postojanje različitih tipova adrenergičkih receptora prvi je predložio Ahlquist (1948). Ovaj autor se oslanjao na razlike u fiziološkim odgovorima na adrenalin, norepinefrin i druge srodne supstance. Bilo je poznato da ovi agensi mogu, ovisno o dozi, organu i specifičnoj tvari, uzrokovati i kontrakciju i opuštanje glatkih mišića. Dakle, norepinefrin na njih ima snažan stimulativni učinak, ali slab - inhibitorni, a izoprenalin - naprotiv; adrenalin ima oba efekta. U tom smislu, Ahlquist je predložio korištenje oznaka a i β za receptore, čija aktivacija dovodi do kontrakcije, odnosno opuštanja glatkih mišića. Izuzetak su glatki mišići gastrointestinalnog trakta - aktivacija oba tipa receptora obično uzrokuje njihovo opuštanje. Aktivnost adrenergičkih stimulansa u odnosu na β-adrenergičke receptore opada u seriji izoprenalin > adrenalin norepinefrin, a u odnosu na α-adrenergičke receptore - u seriji adrenalin > norepinefrin » izoprenalin (tabela 6.3). Ova klasifikacija je potvrđena činjenicom da neki blokatori (na primjer, fenoksibenzamin) eliminiraju utjecaj simpatičkih živaca i adrenergičkih stimulansa samo na α-adrenergičke receptore, a drugi (na primjer, propranolol) - na β-adrenergičke receptore.
β-adrenergički receptori su kasnije podijeljeni na β1 (posebno u miokardu) i β2 (u glatkim mišićima i većini drugih ćelija) podtipove. Ovo se zasnivalo na činjenici da epinefrin i norepinefrin djeluju slično na β1-adrenergičke receptore, ali je epinefrin 10-50 puta jači na β2-adrenergičke receptore (Lands et al., 1967). Razvijeni su selektivni blokatori β1- i β2-adrenergičkih receptora (poglavlje 10). Nakon toga, izolovan je gen koji kodira treći podtip β-adrenergičkih receptora, β3 (Emorine et al., 1989; Granneman et al., 1993). Budući da su β3-adrenergički receptori otprilike 10 puta osjetljiviji na norepinefrin nego na adrenalin, te su relativno otporni na djelovanje blokatora kao što je propranolol, oni mogu biti odgovorni za atipične reakcije nekih organa i tkiva na kateholamine. Takva tkiva uključuju, posebno, masno tkivo. Istovremeno, uloga β3-adrenergičkih receptora u regulaciji lipolize kod ljudi još nije jasna (Rosenbaum et al., 1993; Kriefctal., 1993; Lonnqvist et al., 1993). Pretpostavlja se da polimorfizmi u genu za ovaj receptor mogu biti povezani sa osjetljivošću na gojaznost ili insulin-zavisni dijabetes melitus u nekim populacijama (Armen i HofTstedt, 1999). Zanimljiva je mogućnost upotrebe selektivnih β3-blokatora u liječenju ovih bolesti (Weyer et al., 1999).
Alfa adrenergički receptori se također dijele na podtipove. Prva osnova za ovu podjelu bio je nalaz da norepinefrin i drugi α-adrenergički stimulansi mogu akutno potisnuti oslobađanje norepinefrina iz neurona (Starke, 1987; vidjeti također sliku 6.4). Naprotiv, neki α-blokatori dovode do značajnog povećanja količine noradrenalina koji se oslobađa kada su stimulirani simpatički živci. Pokazalo se da ovaj mehanizam supresije oslobađanja norepinefrina po principu negativne povratne sprege posreduju α-adrenergički receptori, koji se po svojim farmakološkim svojstvima razlikuju od onih koji se nalaze na efektornim organima. Ovi presinaptički adrenergički receptori su nazvani a2, a klasični postsinaptički adrenergički receptori su nazvani a (Langer, 1997). Klonidin i neki drugi adrenergički stimulansi jače djeluju na α2-adrenergičke receptore, a na primjer, fenilefrin i metoksamin - na α1-adrenergičke receptore. Malo je podataka o prisustvu presinaptičkih α1-adrenergičkih receptora u neuronima autonomnog nervnog sistema. Istovremeno, a2-adrenergički receptori su pronađeni u mnogim tkivima i na postsinaptičkim strukturama, pa čak i izvan sinapsi. Dakle, aktivacija postsinaptičkih α2-adrenergičkih receptora u mozgu dovodi do smanjenja tonusa simpatikusa i, očigledno, u velikoj mjeri određuje hipotenzivni učinak klonidina i sličnih lijekova (poglavlje 10). U tom smislu, ideje o isključivo presinaptičkim α2-adrenergičkim receptorima i postsinaptičkim α1-adrenergičkim receptorima treba smatrati zastarjelim (Tabela 6.3).

Tabela 6.5. Podgrupe adrenergičkih receptora
Koristeći metode molekularnog kloniranja, identifikovano je još nekoliko podgrupa unutar oba podtipa α-adrenergičkih receptora (Bylund, 1992). Otkrivene su tri podgrupe α-adrenergičkih receptora (a1A, a1B i a1D; tabela 6.5), koje se razlikuju po farmakološkim svojstvima, strukturi i distribuciji u organizmu. Istovremeno, njihove funkcionalne karakteristike gotovo se ne proučavaju. Među a2-adrenergičkim receptorima, također su izdvojene 3 podgrupe a2B i a2C; sto 6.5), koji se razlikuju u distribuciji u mozgu. Moguće je da barem α2A adrenergički receptori mogu igrati ulogu presinaptičkih autoreceptora (Aantaa et al., 1995; Lakhlani et al., 1997).
Molekularne osnove funkcionisanja adrenergičkih receptora
Očigledno, reakcije na aktivaciju svih vrsta adrenergičkih receptora su posredovane G-proteinima, uzrokujući stvaranje sekundarnih glasnika ili promjene u permeabilnosti jonskih kanala. Kao što je već diskutovano u pogl. 2, takvi sistemi uključuju 3 glavne proteinske komponente - receptor, G protein i efektorski enzim ili kanal. Biohemijske posledice aktivacije adrenergičkih receptora su na mnogo načina iste kao kod M-holinergičkih receptora (vidi gore i tabelu 6.4).
Struktura adrenergičkih receptora
Adrenergički receptori su porodica srodnih proteina. Osim toga, oni su strukturno i funkcionalno
U centralnom nervnom sistemu najveća koncentracija noradrenergičkih neurona nalazi se u locus coeruleusu sive materije ponsa. Odavde se aksoni neurona projektuju na cerebralni korteks, hipokampus, hipotalamus, mali mozak, duguljastu moždinu i kičmenu moždinu.
Adrenergički sistem je prvenstveno povezan sa stimulativnim efektima na funkcije centralnog nervnog sistema.
Dopaminergički sistem
Dopaminski sistemi (krugovi) mozga sisara su dobro proučeni.
Dopaminergički neuroni kod sisara nalaze se prvenstveno u srednjem mozgu (nigro-neostrijatalni sistem) i u hipotalamskom regionu.
Postoje 3 glavna dopaminergička sistema (krugovi):
Prvo. Tijelo neurona nalazi se u hipotalamusu i šalje kratki akson u hipofizu.
Ovaj put je dio hipotalamus-hipofiznog sistema i kontrolira sistem endokrinih žlijezda.
Sekunda. Ovo substantia nigra . Aksoni ovih neurona projektuju se na striatum.
Ovaj sistem sadrži ¾ moždanog dopamina. Ključan je u regulaciji toničnih pokreta.
Nedostatak dopamina u ovom sistemu dovodi do Parkinsonove bolesti. U ovom slučaju, neuroni crne supstance umiru. Primjena L-DOPA (prekursora dopamina) ublažava neke simptome bolesti kod pacijenata.
Treće. Tijela neurona leže u srednjem mozgu pored crne supstance, a aksoni se projektuju na preliježe moždane strukture, korteks, limbički sistem, posebno frontalni korteks, septalni dio i entorhinalni korteks. Entorhinalni korteks je glavni izvor projekcija do hipokampusa.
Struktura ovog dopaminergičkog sistema je dobro shvaćena, ali je njegova funkcija slabo shvaćena. Ovaj sistem je veoma aktivan kod šizofrenije. Kada se potisne (hlorpromazin, haloperidol), potiskuju se neki simptomi šizofrenije.
Dopaminergički sistem se reguliše kroz različite tipove dopaminskih (D‑) post- i presinaptičkih receptora. Stimulacija presinaptičkih dopaminskih receptora smanjuje sintezu i oslobađanje dopamina iz nervnih završetaka.
Postoje dvije grupe dopaminskih receptora: grupa D 1 receptora (podgrupe D 1 i D 5) uglavnom uzrokuje postsinaptičku inhibiciju. Oni su povezani sa Gs proteinima. Stimulirati adenilat ciklazu, povećavajući sadržaj cAMP. Grupa D 2 receptora (podgrupe D2 -, D3 - , i D4 - ) uzrokuje pre- i postsinaptičku inhibiciju. Ovi receptori su povezani sa Gi/o proteinima. Inhibira adenilat ciklazu. Osim toga, aktiviraju K+ kanale i imaju inhibitorni učinak na Ca 2+ kanale.
Serotonergički sistem
Važno mjesto među CNS medijatorima/modulatorima pripada serotoninu (5‑hidroksitriptamin; 5-HT). Najobimnija zbirka serotonergičkih neurona nalazi se u gornjem dijelu produžene moždine i u mostu. Ove formacije se nazivaju raphe nuclei (nucleus raphes). Njihovi neuroni projiciraju i kranijalno (korteks, hipokampus, limbički sistem, hipotalamus) i kaudalno (dužna moždina i kičmena moždina). Najveći sadržaj serotoninskih receptora nalazi se u hipokampusu, strijatumu i frontalnom korteksu.
Ekscitacija presinaptičkih receptora smanjuje oslobađanje serotonina i nekih drugih transmitera iz nervnih završetaka. Što se tiče postsinaptičkih receptora, njihova stimulacija može biti praćena i ekscitacijom i inhibicijom.
Postoji 7 podtipova serotoninskih receptora sa dodatnim podjelama za pojedinačne podtipove (5-HT 1 A - D, 5-HT 2 A - C, itd.). Za funkcije centralnog nervnog sistema uočava se važna uloga prva četiri podtipa. Fiziološki značaj posljednja tri (klonirana) podtipa nije poznat.
5-HT1 receptori su lokalizovani pre- i postsinaptički. Dakle, stimulacija 5-HT1A receptora uzrokuje postsinaptičku inhibiciju. Presinaptička inhibicija je povezana s funkcijom 5‑HT1D receptora. Prenos postsinaptičke ekscitacije povezan je sa 5-HT 1 C, 5-HT 12, 5-HT 3 i 5-HT 4 receptorima.
Funkcija serotonergičkog sistema je prilično raznolika. To je regulacija ciklusa spavanja i budnosti, mentalnih funkcija, raspoloženja, pamćenja, apetita, ekscitabilnosti motornih neurona, regulacije provođenja senzornih nadražaja (uključujući bol), centralne termoregulacije, utjecaja na proizvodnju niza hipotalamičkih faktora i hormoni hipofize.
Poznati su lekovi koji utiču na serotonergički sistem.
Acetilholinergički sistem
Funkcija holinergičkih receptora u centralnom nervnom sistemu nije dovoljno jasna (posebno N-holinergičkih receptora). Poznato je da kolinergički procesi učestvuju u kontroli mentalnih i motoričkih funkcija, u reakciji buđenja i u učenju.
Acetilholin stupa u interakciju s M- i H-holinergičkim receptorima koji se nalaze u različitim dijelovima mozga i moždanog stabla.
Holinergički receptori su lokalizovani i post- i presinaptički.
U pravilu, acetilholin djeluje kao ekscitatorni neurotransmiter. U nekim slučajevima dolazi do efekta kočenja.
Ekscitacija presinaptičkih M-holinergičkih receptora smanjuje oslobađanje acetilholina.
U medicinskoj praksi centralni antiholinergički blokatori se koriste kao anksiolitici (Amizil) i za parkinsonizam (Cyclodol). Posljednjih godina su supstance koje aktiviraju centralne kolinergičke procese (na primjer, antiholinesterazni lijekovi koji lako prodiru kroz krvno-moždanu barijeru, uključujući fizostigmin) privukli veliku pažnju. To je zbog činjenice da u nekim slučajevima imaju blagotvoran učinak kod Alchajmerove bolesti (presenilna demencija), kod koje je smanjen sadržaj kolinergičkih neurona u mozgu.
Medijatorski sistemi aminokiselina
Medijatori uključuju γ-aminobuternu kiselinu (GABA), glicin i, očigledno, glutamat. Osim toga, pretpostavlja se da brojne druge aminokiseline mogu biti neurotransmiteri ili neuromodulatori (L-aspartat, β-alanin, itd.).
Adrenalin je prvi put otkriven u ekstraktima nadbubrežne žlijezde 1895. godine. Godine 1901. izvršena je sinteza kristalnog adrenalina. Ubrzo je adrenalin pronašao upotrebu u medicini za povećanje krvnog pritiska tokom kolapsa, za sužavanje krvnih sudova tokom lokalne anestezije, a zatim i za ublažavanje napada bronhijalne astme. Godine 1905 otkriven je važan fiziološki značaj adrenalina. Na osnovu sličnosti djelovanja adrenalina sa efektima uočenim tijekom iritacije simpatičkih nervnih vlakana, sugerirano je da se prijenos nervnog pobuđenja sa simpatičkih nervnih završetaka na efektorske ćelije vrši uz učešće hemijskog transmitera (medijatora), što je adrenalin ili supstance slične adrenolinu. To je označilo početak doktrine o hemijskom prijenosu nervnog uzbuđenja. Potom je otkriven proces biosinteze adrenalina, počevši od aminokiseline tirozina, preko dioksifenilalanina (L-dopa), dopamina, norepinefrina do adrenalina. Godine 1946 utvrđeno je da glavni medijator adrenergičke (simpatičke) transmisije nije sam adrenalin, već norepinefrin. Endogeni adrenalin koji se formira u tijelu djelomično je uključen u procese nervne stimulacije, ali uglavnom igra ulogu hormonske supstance koja utječe na metaboličke procese. Norepinefrin ima posredničku funkciju u perifernim nervnim završecima i u sinapsama centralnog nervnog sistema. Biohemijski sistemi tkiva koji stupaju u interakciju sa norepinefrinom nazivaju se adrenoreaktivni (adrenergički) sistemi ili adrenoreceptori („adrenoreceptori“). Prema modernim shvatanjima, norepinefrin, koji se oslobađa tokom nervnog impulsa iz presinaptičkih nervnih završetaka, utiče na norepinefrin osetljivu adenilat ciklazu stanične membrane adrenoreceptorskog sistema, što dovodi do povećanog stvaranja unutarćelijskog 3"-5"-cikličkog adenozin monofosfata (c. ), koji igra ulogu „sekundarnog“ transmitera (medijatora), za aktiviranje biosinteze visokoenergetskih jedinjenja i dalje za sprovođenje adrenergičkih fizioloških efekata. Dopamin, koji je hemijski prekursor norepinefrina, ali ima nezavisnu ulogu neurotransmitera, takođe igra važnu ulogu u prenosu impulsa do centralnog nervnog sistema.
Formiranje medijatora se pretpostavlja prema sljedećoj shemi: fenilalanin -> tirozin -> dioksifenilalanin (DOPA) -> dopamin (1. medijator, kateholamin) -> norepinefrin (glavna uloga u prijenosu ekscitacije u adrenergičkim sinapsama). Norepinefrin u sinapsama i nadbubrežnim žlijezdama može se pretvoriti u adrenalin i obrnuto).
Počevši od treće reakcije, javlja se u nervnim ćelijama (prva reakcija se javlja u jetri). Neurotransmiteri se spuštaju duž aksona u vezikulama do presimpatičkih završetaka. Joni magnezija učestvuju u procesu transporta vezikula. Medijatori mogu biti uništeni MAO tipa A (uništavaju norepinefrin, adrenolin i serotonin). Za zaštitu od MAO, norepinefrin i adrenalin se kombinuju sa posebnim proteinima i ATP-om (formira se depo). To su stabilne granule (stabilna frakcija). Labilna frakcija je predstavljena nevezanim medijatorom u vezikulama. Osim toga, u citoplazmi postoji mala količina slobodnog adrenalina, ali se lako uništava enzimima.
Nakon otpuštanja transmitera u sinaptičku pukotinu, njegov višak može biti uništen pomoću COMT-a. Može doći i do ponovnog preuzimanja dijela transmitera presinaptičkom membranom.
Utjecaj adrenalina na krvni tlak uključuje nekoliko faza: u prvoj fazi se aktiviraju β1-adrenergički receptori miokarda, što dovodi do povećanja minutnog volumena; u drugom - kašnjenje u porastu pritiska (vagodepresorski refleksni efekat); treća faza je praćena uticajem adrenalina na b (porast) i c (smanjenje) vaskularnih receptora i četvrta - hipotenzija u tragovima, brzi neuronski unos adrenalina, inaktivacija njegovog viška enzimom COMT.
U regulaciji tjelesnih funkcija, pored klasičnih medijatora, važnu ulogu imaju regulatorni faktori peptidne prirode. Regulatorni peptidi su široko rasprostranjeni u različitim tkivima, uključujući i nervno tkivo. Oni učestvuju u neurohemijskim mehanizmima koji podržavaju osnovne homeostatske konstante tela, formiraju i provode ciljno usmereno ponašanje, kao i u procesima koji kontrolišu emocionalnu sferu, motivaciju i pamćenje. Vjerovatno je da biološki aktivni peptidi igraju važnu ulogu u integraciji funkcionalnih sistema tijela, osiguravajući njihov koordiniran rad u promjenjivim uvjetima okoline. Imaju ključnu ulogu u regulaciji imunološke odbrane, u pokretanju adaptivnih odbrambenih reakcija tokom infekcije, oštećenja tkiva, stresa, kao i u formiranju patoloških stanja u organizmu, uključujući alkoholizam. Mnogi neuropeptidi su uključeni u regulaciju starosnih promjena, uključujući procese puberteta.
Jedna od faza metabolizma peptida je ograničena proteoliza, koja igra glavnu ulogu kako u procesima njihove biosinteze tako iu procesima inaktivacije. Peptidne hidrolaze, koje obrađuju i razgrađuju peptidne regulatore, osiguravaju njihovo funkcioniranje i određeni odnos u tijelu.
Većina neuropeptidnih prekursora uključuje peptidne sekvence s različitim biološkim aktivnostima. Koji će se tačno peptidi formirati iz prekursora ovisi o skupu proteinaza koje djeluju na molekul prekursora i o odnosu njihovih aktivnosti.
Sa parakrinim djelovanjem peptida, aktivnost ekstracelularnih peptidaza određuje životni vijek peptida, udaljenost na kojoj se može difundirati i, posljedično, raspon ciljeva na koje djeluje. Tako se uz pomoć proteinaza regulišu fiziološki efekti peptida u fazi biosinteze i u fazi inaktivacije peptida.
Posebnost peptidne regulacije funkcionalnog stanja organizma je da se u svakom trenutku mora održavati potrebna koncentracija određenih peptida u svakom području. To se može postići preciznim i koordiniranim radom proteinaza koje vrše sintezu i razgradnju peptida, odnosno održavanjem određenog prostorno-temporalnog mozaika proteolitičke aktivnosti u mozgu. Kada se spoljni uslovi promene, ili pod bilo kojim uticajem (npr. alkoholizam), ovaj mozaik se menja na određeni način kako bi se obezbedilo funkcionisanje funkcionalnih sistema organizma u novim uslovima.
Završna faza formiranja aktivnih peptida iz neaktivnih prekursora i početne faze njihove degradacije uključuju bazične karboksipeptidaze - enzime koji cijepaju osnovne aminokiselinske ostatke (arginin i lizin) sa C-kraja peptida. To uključuje, posebno, karboksipeptidazu H i nedavno otkrivenu karboksipeptidazu inhibiranu PMSF-om. Oni igraju važnu ulogu u regulaciji nivoa aktivnih neuropeptida u organizmu, što objašnjava interesovanje za proučavanje ovih enzima, uključujući različite fiziološke i patološke procese koji se dešavaju u telu.
Djelovanje adrenergičkih blokatora prvenstveno je usmjereno na b, b-adrenergičke receptore. Kada djeluju na β1-adrenergičke receptore, joni kalcija počinju da ulaze u ćeliju, djelujući direktno stimulativno. Osim toga, aktivira se fosfolipaza C. Ona razlaže membranski fosfolipid na dvije aktivne tvari: inozitol-3-fosfat, koji stimulira oslobađanje kalcija iz intracelularnih zaliha u citoplazmi, i diacilglicerol, koji aktivira protein kinaze. Protein kinaze aktiviraju fosforilaze, koje fosforiliraju proteine. Djelujući na β-receptore preko regulatornog proteina Gs, aktivira se adenilat ciklaza, a produkt njenog rada cAMP aktivira protein kinaze. Kada djeluje na β2 receptore preko Gi proteina, adenilat ciklaza se inhibira. I G i Gi zahtijevaju GTP za svoju funkciju.
Konkretno, beta-blokatori, dok ispoljavaju presorski efekat, karakterišu prisustvo nuspojava, kao što su arterijska hipotenzija, bradikardija itd., koje je teško objasniti isključivo dejstvom ovog leka na receptore. Možda su neki od efekata posredovani peptidergijskim sistemom, jer promjene u adrenergičkom sistemu uzrokuju promjene u nivou regulatornih peptida: vazopresina, angeotenezina i samatropina.
Također preporučujemo
 Racionalno korištenje i zaštita vodnih resursa - Hipermarket znanja Izvori i korištenje slatke vode
Racionalno korištenje i zaštita vodnih resursa - Hipermarket znanja Izvori i korištenje slatke vode
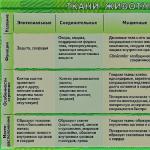 Vrste biljnih tkiva i njihove funkcije
Vrste biljnih tkiva i njihove funkcije
 Građa listova biljaka, tipovi rasporeda listova, fotosinteza i transpiracija
Građa listova biljaka, tipovi rasporeda listova, fotosinteza i transpiracija
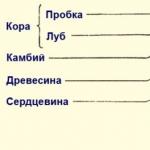 Funkcije i struktura stabljike biljke
Funkcije i struktura stabljike biljke
 Recept za džem od pomorandže sa koricom sa fotografijom
Recept za džem od pomorandže sa koricom sa fotografijom
 Posna jela od bundeve - zdrava sorta
Posna jela od bundeve - zdrava sorta